I. 서 론
II. 향고래 클릭 신호 측정
III. 분석 및 결과
IV. 결론 및 고찰
I. 서 론
향고래(Physeter macrocephalus)는 이빨고래 중 가장 크며, 포유류 중에서 인간과 범고래를 제외하고는 가장 넓은 지역에 분포한다.[1,2,3] 일반적으로 수심이 깊은 해역에 서식하며, 특히 용승으로 인해 1차 생산성이 높은 지역에서 주로 발견된다.[2,3] 과거의 포경, 혼획 및 좌초 자료에 의하면 국내의 향고래 발견 은 매년 1 ~ 2회 정도로 보고되고 있다.[4]
향고래 머리는 거대한 두개골과 비강 복합체(nasal complex)로 이루어져 있다(Fig. 1). 아래턱 위와 두개골 앞에 위치한 비강 복합체는 전체 길이의 1/4에서 1/3을 차지하며, 경랍(spermaceti), 멜론(junk or melon), 기낭(air sacs), 비강(nasal passages)으로 구성되어 있다.[3,5,6]그 중 비강은 비대칭으로 왼쪽은 호흡하는데 사용하며, 오른쪽은 소리를 생성하는데 특화되어 있다.[5] 향고래는 일반적으로 클릭으로 정의되는 짧은 음향 신호를 연속적으로 발생시켜 환경을 탐색하고 동종과 의사소통하는 것으로 알려져 있으며, 이 클릭 신호는 향고래의 독특한 머리 구조로 인해 다중 펄스 구조를 가진다.[5,7,9]
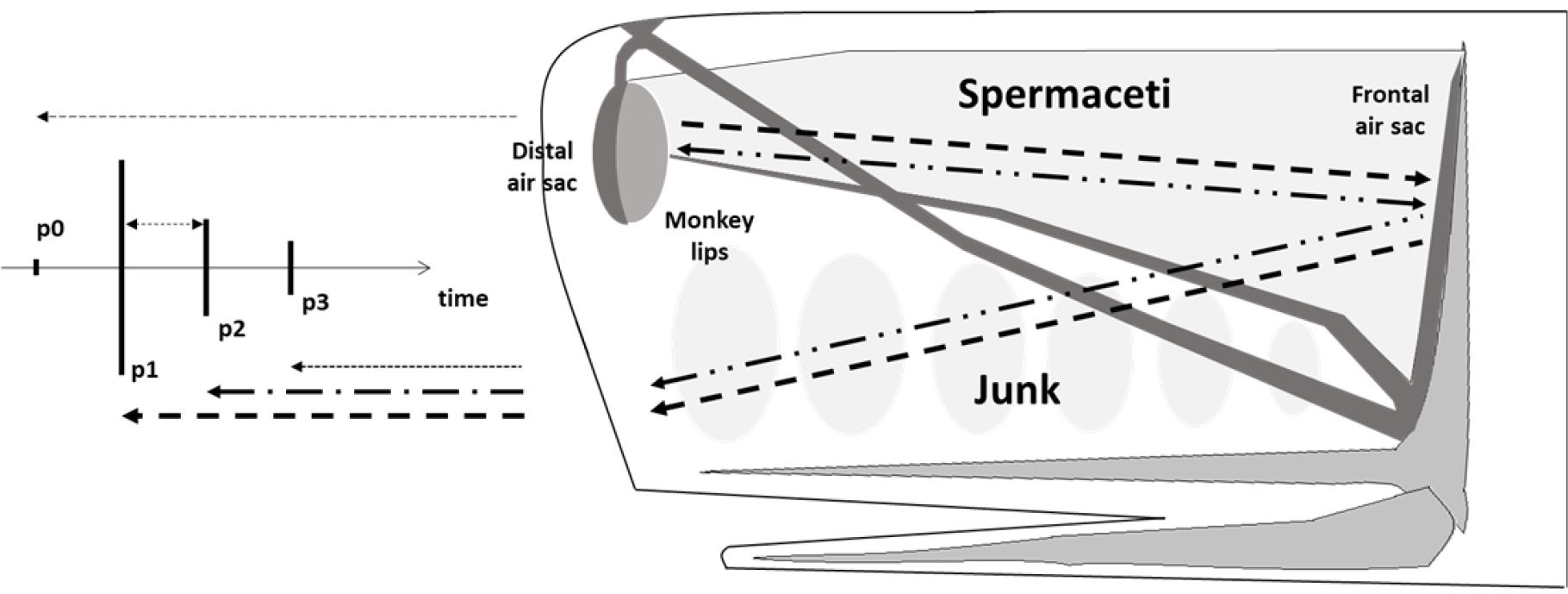
Fig. 1
A scheme of bent-horn model of sound production of a click in sperm whales’s head [modified from Fig. 1 of Caruso et al.[5]].
두개골 앞에 있는 떨림판(monkey lips or museau de singe)에서 생성된 소리는 매우 약한 신호만이 원위 기낭(distal air sac)을 통과하여 정면으로 방출되며, 이 신호를 본 논문에서는 p0로 정의한다. p0로 방출되지 않은 대부분의 소리 에너지는 원위 기낭에서 반사되어 경랍 기관을 따라 전파된 후 전두 기낭(frontal air sac)에서 반사되어 멜론을 통해 앞으로 이동하여 하부에서 방출된다. 이 신호는 첫 번째 신호에 비해 강한 에너지를 가지며 본 논문에서는 p1으로 정의한다.[10,11,12] 전두 기낭에 반사된 신호 중 일부는 다시 경랍 기관으로 이동하여, 두 기낭 사이에서 앞뒤로 반사되며 멜론을 통해 기하급수적으로 감소된 진폭으로 신호를 방출하면서 p2, p3 신호들을 연속적으로 생성한다.[5,13] p1을 포함한 그 이후의 신호들은 동일한 위치인 멜론에서 방출되고 동일한 전파 경로를 가지므로, p1–p2, p2–p3 같이 펄스 사이의 시간 간격(Inter-Pulse Interval, IPI)은 모든 클릭에 대해 동일하다.[5,14]
측정된 향고래의 클릭으로부터 IPI를 계산하기 위해 다양한 신호처리 기술과 분류 기준을 적용하였으나,[15,16,17,18,19] 과거에는 다중 펄스 구조가 명확하게 나타나지 않는 다수의 클릭은 분석에 적합하지 않은 것으로 간주되어 제거되고 소수의 클릭만 선택하여 IPI를 계산하였다.[20] 이후 Zimmer et al.[12]에 의해 off-axis에서는 전두 기낭에서 반사된 신호가 멜론을 통하지 않고 바로 외부로 빠져나가는 p1/2 신호가 발생함을 보고하였다. p1/2 신호는 고래와 수신기의 각도에 따라 p0와 p1사이에서 발생하며, 이 신호로 인해 다중 펄스 형태가 명확하게 나타나지 않을 수 있다. 따라서 향고래 IPI에 대한 이전 연구들에서 제거한 클릭은 off-axis에서 수신된 신호일 가능성이 높다.[12,20] Teloni et al.[14]는 bent horn model을 이용하여 IPI가 항상 일정한 간격으로 발생되므로, 각 개체가 발생시키는 수백 번의 클릭으로 켑스트럼 분석한 결과를 앙상블 평균하면 IPI가 일관되게 추정된다는 것을 보였다. 또한 Antunes et al.[21]은 자기상관(autocorrelation)에 기반한 방법이 IPI 계산에 적합함을 보였다.
IPI와 향고래 전장의 상관성에 대한 연구가 다수 진행되었다. Norris와 Harvey[8]는 향고래의 클릭 신호 발생 이론이 정립되기 전부터 하나의 펄스가 고래의 머리 내에서 반사되어 다중 펄스 구조가 발생한다고 추측하였다. 따라서 IPI는 소리가 고래의 머리 내에서 이동하는데 걸리는 시간이므로, 머리 길이와 고래 길이 사이의 상대성장 관계(allometric relationships)로 인해 IPI가 고래 전장의 척도로 사용될 수 있다고 가설을 세우고 연구를 진행하였다. Rhinelander와 Dawson[18]은 IPI가 단일 개체별로 일정하지만 전장이 다른 개체 간에 차이가 있음을 밝혀냈으며, Gordon,[16] Growcott et al.[22]은 간단한 사진 측정 기법을 사용하여 추정한 향고래 길이와 IPI 간의 관계를 정량화하여 경험식을 제안하였다. 이 추정 기법은 비침투적(non-invasive) 방법이라는 장점으로 인해 수년에 걸쳐 동일한 개체의 전장을 기록함으로써 성장을 정량화하거나 개체군 구조 추정과 같은 전 세계에 서식하는 향고래의 생태 역학(ecological dynamics) 모니터링에 사용되고 있다.[8,9,14,15,16,17,18,19,20,21,22,23,24,25,26]
향고래는 성별에 따라 크기나 무게가 다른 이형적인 종이며, 생물학적 측면과 마찬가지로 분포 특성 또한 매우 다르다.[3] 향고래는 한반도 주변에서 자주 발견되는 종이 아니기 때문에 국내에서의 향고래 연구는 거의 진행되지 않아, 동해에서 발견되는 향고래에 대한 정보는 부족한 현실이다. 본 연구에서는 동해에서 획득한 향고래의 클릭 신호에서 IPI를 계산하고, 향고래 전장과의 관계식에 대입하여 향고래의 크기를 추정하였다. 이는 국내에서 음향을 이용하여 해양포유류의 크기를 추정하는 최초의 시도라는데 의의가 있다. 또한 음향학적으로 추정된 향고래의 크기를 통해 동해에서 발견되는 향고래의 생태학적 특징을 파악하였으며, 향고래의 분포 특성 연구를 위한 기초자료로 제공될 수 있다.
II. 향고래 클릭 신호 측정
향고래의 음향 신호는 국립수산과학원 고래연구센터에서 2017년 10월 25일부터 11월 8일까지 실시한 2017년도 제3차 동해 소형고래류 조사에서 획득하였다. 2017년 10월 27일 직선횡단 목시 조사 중 포항 앞바다(35° 0' 6.12" N, 129° 44' 28.26" E, 총 수심 120 m)에서 향고래 6마리를 발견하였으며, 목시 관측을 위해 약 30 min간 8.8 km를 이동하며 향고래 무리를 추적하였다(Fig. 2). 음향 녹음은 목시 관측이 종료된 이후 시작되었으며, 최초 발견 지점에서 약 4.3 km 떨어진 위치(35° 58' 7.62" N, 129° 45' 59.64" E)에서 조사선을 정지시켜 수동 탐지방법으로 향고래가 발생시키는 신호를 녹음하였다. 수중청음기(SM3BAT/H1, Wildlife Acoustics, Inc., USA)는 선박 측면에서 수심 5 m에 위치시켰으며, 샘플링주파수 384 kHz로 설정 후 약 6 min간 측정하였다.
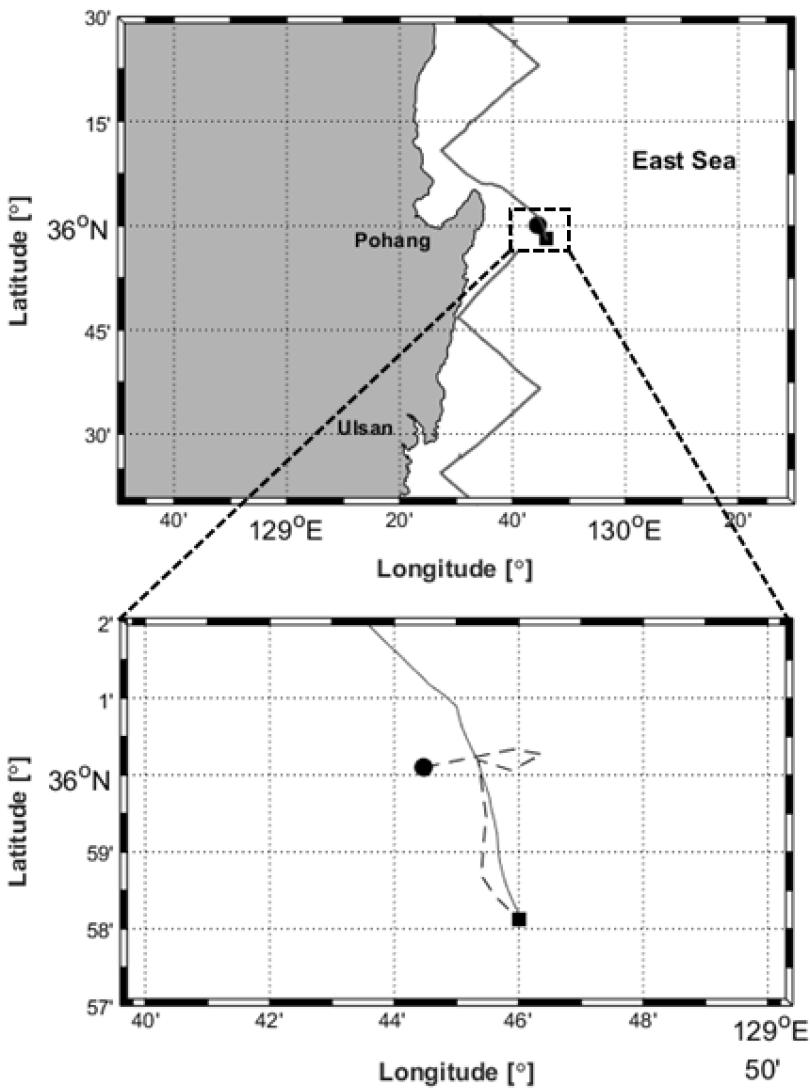
Fig. 2
Measurements area for recording of sperm whale(black square), and first contact position(black circle) in the East Sea. The line transect sighting survey and tracking whales are shown as gray solid lines and black dashed line, respectively.
III. 분석 및 결과
수신된 향고래의 클릭 신호는 다중 펄스 구조의 규칙적인 시퀀스(sequence)로 발생되는 클릭 트레인(click trains) 형태를 보였다(Fig. 3). 일반적으로 향고래의 클릭 신호는 3 kHz ~ 16 kHz 대역에서 우세한 주파수 특성을 가지므로,[5,14] 그 대역폭에 해당하는 Finite Impulse Response(FIR) 대역통과 필터를 적용하여 분석을 수행하였다. 필터된 신호 중 클릭 신호를 추출하기 위해 피크 신호 대 잡음비[peak SNR(Signal to Noise Ratio)]가 문턱값을 초과하는 신호를 선택하는 문턱값 탐지기(threshold detector)를 이용하였다.[27] 즉, 설정 문턱값보다 낮은 신호는 소음으로 간주되며, 설정 문턱값 이상의 모든 과도 신호(transient signal)는 향고래의 클릭 신호로 판단한다. 이때 설정 문턱값은 배경소음보다 높아야 하며, 기존의 향고래 크기 추정 연구 논문들은 수신된 클릭의 크기나 측정 환경에 따라 적절히 설정하였다. 본 연구에서는 수신된 전체 시계열 신호로 배경소음을 계산하고 배경소음보다 23.5 dB 높은 값으로 문턱값을 설정하여 클릭 신호를 추출하였다.
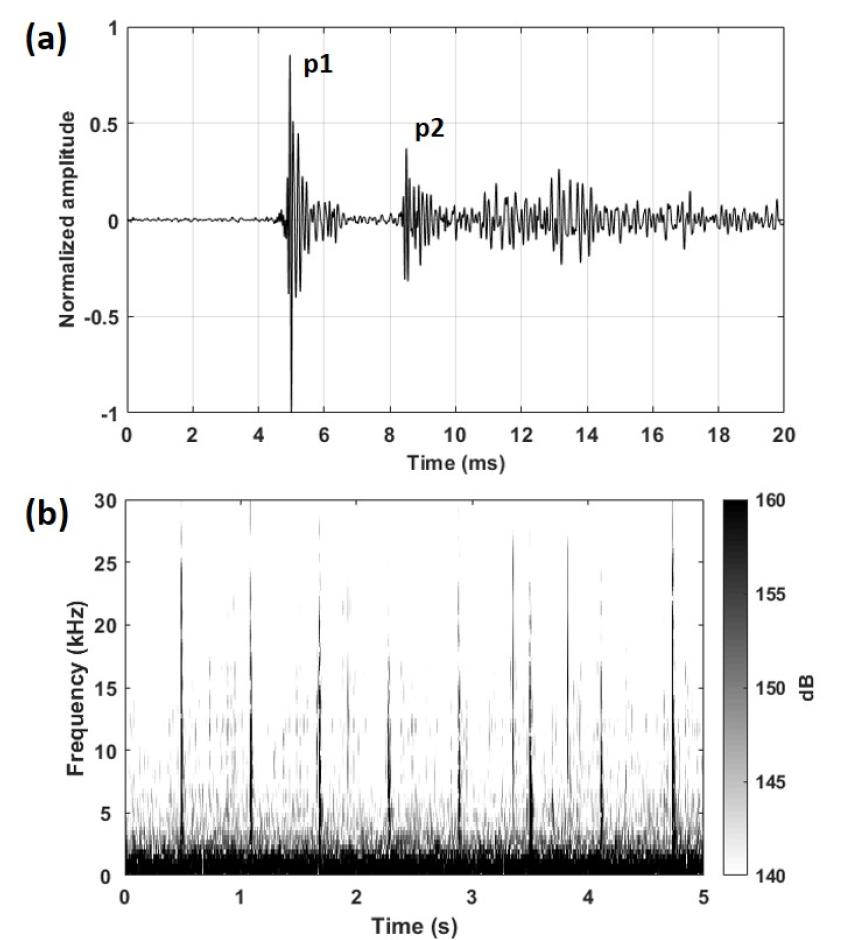
Fig. 3
(a) Waveform of a sperm whale click showing multi-pulse structure, (b) Spectrogram of clicks produced by sperm whale.
추출된 클릭은 개체 간의 간섭을 제거하기 위해, 동일한 개체에서 발생되는 클릭의 시간적 분리를 정량화하는 클릭 간 간격(Inter Click Interval, ICI)을 고려하여 선택하였다. 일반적으로 동일한 개체에서 발생된 클릭은 개별 활동에 의해 규칙적인 시퀀스를 가지므로 짧은 시간 내에서의 ICI는 크게 변하지 않는다.[27] ICI의 순간적인 변화는 다른 개체에 의해 발생하는 경향을 가지므로, 연속적인 ICI를 가지지 않는 클릭들은 제외하고 분석을 실시하였다(Fig. 4).
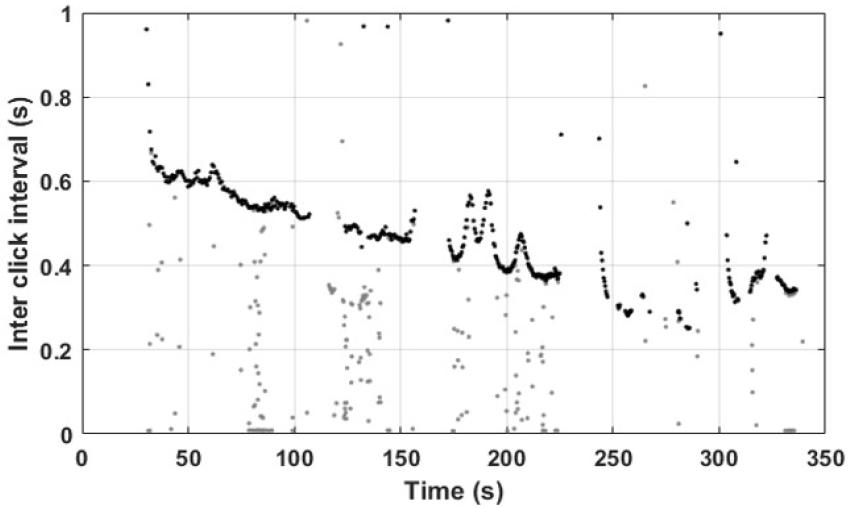
Fig. 4
Inter Click Interval (ICI) of total clicks (gray dots) and selected clicks for computing IPI (black dots).
클릭의 다중 펄스 구조는 향고래와 수신기의 기하학적 위치에 따라 명확하게 나타나지 않는 경우가 많다.[12] 최근에는 수신기 배열을 통해 다중 펄스 구조가 명확하게 나타나는 축 상의 신호만 선택하여 계산하는 방법도 많이 사용되지만, 본 연구에서는 단일 수신기만으로 신호를 수신하였으며 향고래의 위치나 자세각 등의 추가적인 정보 또한 부족하였다. 따라서 IPI 측정은 Teloni et al.[14]가 제안한 켑스트럼 결과를 앙상블 평균하는 방법을 이용하였다. 이 방법은 고래의 off-axis에서 발생시키는 p1/2 신호나 해수면 반사파와 같은 간섭에 의한 영향을 줄여줄 수 있으므로 기존 방법들이 분석에 사용하지 않았던 다중 펄스 구조가 명확하게 나타나지 않는 클릭 신호들도 분석에 사용할 수 있다.[14,20] 각 개별 클릭의 켑스트럼은 다음과 같이 계산된다.
여기서 는 향고래 클릭의 시계열 신호이며, 및 은 각각 고속 푸리에 변환 및 역 고속 푸리에 변환이다.[14,28] 켑스트럼은 파워 스펙트럼의 로그 값을 시간 축으로 다시 변환한 결과이며, 피크 값은 시간축의 신호()에서 반복되는 구조(펄스) 사이의 시간 간격과 동일하게 나타난다. 켑스트럼 결과에서 피크가 나타나려면 분석에 사용되는 신호의 길이는 펄스 간 간격보다 적어도 두 배 이상 길어야하므로,[14] 본 논문에서는 각각의 클릭을 20 ms의 시간으로 추출하여 분석을 실시하였다.
약 6 s간 측정된 신호 중 선택된 모든 클릭에 대한 켑스트럼 분석 결과 약 3 ms ~ 4 ms 대역에서 켑스트럼 피크를 나타내었다[Fig. 5(a)]. IPI를 계산하기 위해서는 단일 개체에서 발생시킨 클릭만을 이용하여 켑스트럼 결과를 평균해야 하지만, 음향 조사 전 수행된 목시 조사에서 발견된 향고래는 6마리로 분석에 사용된 클릭은 몇 마리의 개체가 발생시켰는지 정확히 알 수 없다. 따라서 클릭 전체가 한 개체의 향고래에서 발생되었다고 가정한 결과와 각각의 클릭 트레인마다 다른 개체로 가정한 결과를 비교하였다. 총 450개 클릭이 한 개체의 향고래에서 발생되었다고 가정하여 모든 클릭의 켑스트럼 값을 평균한 결과 IPI는 3.94 ms에서 최대 값으로 나타났다[Fig. 5(b)]. 클릭이 10 s 이상 수신되지 않는 경우 다른 개체라고 가정하였을 때 클릭 트레인은 6개로 분리되며(Table 1), 각각의 클릭 트레인의 IPI 범위는 3.51 ms ~ 4.14 ms로 계산되었다. 이때 클릭 수가 10개 미만의 클릭 트레인은 IPI 계산에 포함하지 않았다.
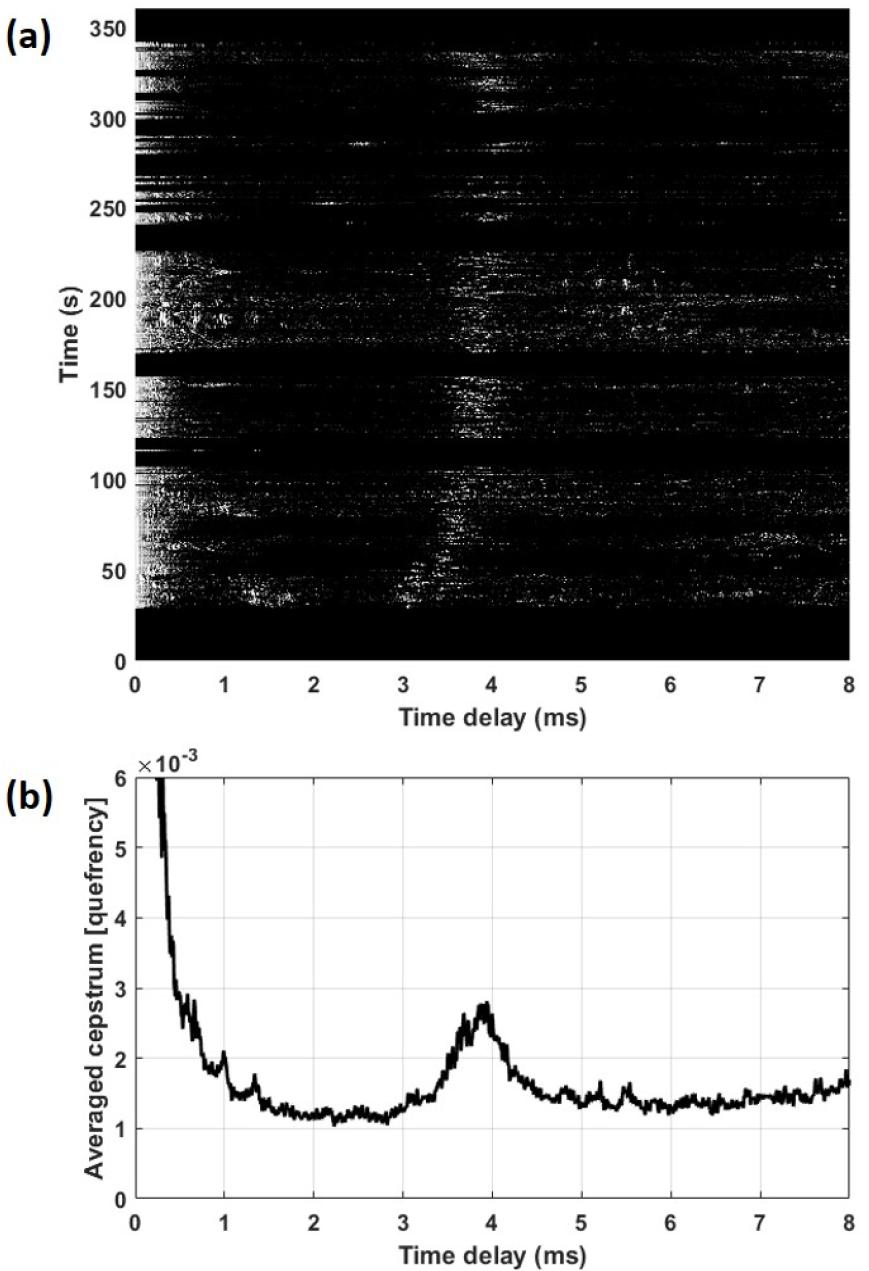
Fig. 5
(a) Cepstrum analysis for 450 clicks, (b) Averaged values of cepstrum.
Table 1.
List of time, number of clicks and IPI for click trains.
|
Time (s)
|
number of clicks
|
IPI (ms)
|
|
29 ~ 107
|
130
|
3.51
|
|
123 ~ 157
|
64
|
3.85
|
|
172 ~ 225
|
123
|
3.80
|
|
241 ~ 267
|
34
|
3.95
|
|
280 ~ 290
|
14
|
4.14
|
|
303 ~ 341
|
69
|
3.90
|
IPI와 향고래 전장의 관계에 대한 회귀방정식은 Gorden[16]이 제안한 Eq. (2)와 Growcott et al.[22]가 제안한 Eq. (3)이 있다.
Gordon[16]은 향고래(대부분 어린 개체)의 사진 측정을 통한 크기 추정을 기반으로 고래의 전장()과 IPI 사이의 경험적 관계를 제안하였으며, 11 m 미만의 개체(IPI < 4 ms)를 측정하는데 적합한 것으로 알려져 있다. 반면에 Growcott et al.[22]은 12 m 이상의 큰 향고래의 IPI와 사진 측정 데이터를 사용하여 공식을 제안하였으며, 11 m 이상의 큰 개체(IPI > 4 ms)의 측정에 적합한 것으로 알려져 있다.
본 연구에서 측정한 동해 향고래의 클릭 신호가 동일 개체에서 발생된 것이라고 가정하였을 때 IPI는 3.94 ms이므로, 위의 두 식에 대입하였을 때 향고래의 전장은 각각 10.54 m와 10.69 m로 나타났다. 또한 수신된 클릭이 여러 개체라고 가정하여 각각의 IPI를 식에 대입해도 전장의 범위는 9.9 m ~ 10.9 m로 추정되었다(Fig. 6).
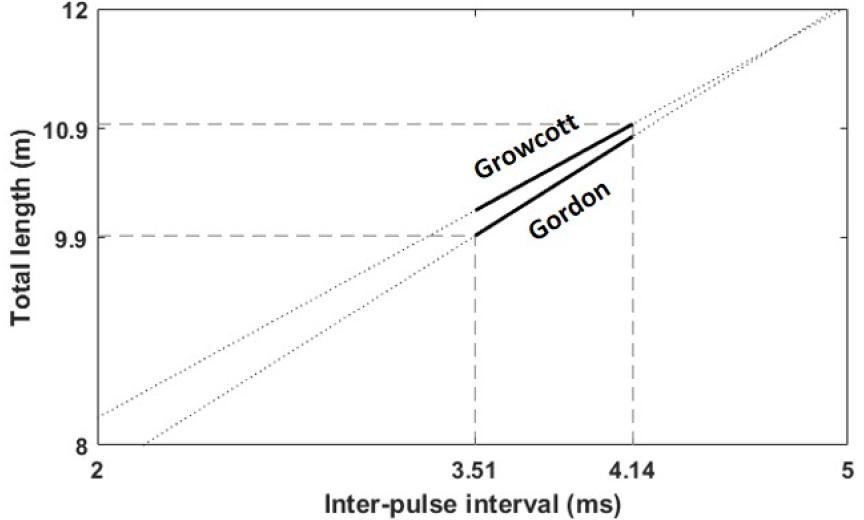
Fig. 6
Range of total length estimates of sperm whale from empirical equation using IPI.
IV. 결론 및 고찰
동해에서 발견된 향고래의 크기를 추정하기 위해 향고래의 클릭 신호를 분석하였다. 향고래의 전장은 클릭을 구성하는 펄스 사이의 시간 간격인 IPI를 이용하여 추정할 수 있다.[29] 본 논문에서는 향고래의 자세, 수심, 거리에 의해 발생하는 효과를 줄일 수 있는 켑스트럼 평균을 이용하여 IPI를 계산하였다. 향고래 전장과 IPI의 관계식에 의해 동해에서 발견된 향고래의 전장은 약 9.9 m ~ 10.9 m로 추정된다. 일반적으로 향고래 수컷은 약 16 m, 암컷은 11 m까지 성장하므로,[3] 본 연구에서 추정한 향고래의 전장은 성체 암컷 또는 유년기 수컷으로 예상할 수 있다. 동해에서 발견된 향고래는 6마리로 구성되어 있는 무리였으며, 일반적으로 향고래 무리는 암컷 향고래들과 그들의 새끼로 구성된 가족 단위로 생활하는 것으로 알려져 있다.[3] 따라서 측정된 신호는 성체 암컷 향고래의 클릭일 가능성이 매우 높으나 이를 뒷받침하기 위한 추가적인 정보는 부족하다.
음향 녹음 시작 직후 향고래 무리는 수중으로 잠수하여, 지속적으로 추적하며 음향신호를 획득하지 못하였다. 또한 발견된 개체수가 많아 하나의 개체가 발생시킨 클릭 트레인이라고 단정 짓기에는 한계가 있었다. 그러나 전체 클릭을 이용하여 추정된 전장과 클릭 트레인 별로 분리하여 추정된 전장의 차이는 1 m 이내로 차이가 크지는 않았다. 따라서 본 연구에서 음향학적으로 추정한 향고래 크기를 다른 지역의 선행 연구 결과와 비교하여 결과의 타당성을 확인하였다. 이오니아해에서 녹음된 향고래 클릭 신호의 켑스트럼 결과를 앙상블 평균하여 계산된 IPI의 범위는 2.1 ms ~ 6.4 ms로 나타났다. 음향학적으로 추정된 향고래의 전장은 7.5 m ~ 14 m로 분포하였으며, 183개의 IPI값이 크기 추정에 사용되었다. 84.7 %의 향고래가 전장이 9 m ~ 12 m로 추정되었으며, 14 m이상의 성체 수컷은 이 지역에서는 발견되지 않았다. 뉴질랜드 남부해역 3개 정점에서 측정된 향고래의 신호는 3.3 ms ~ 10 ms의 IPI 범위를 가지는 것으로 확인되었으며, 997개의 IPI를 이용하여 향고래 크기를 추정하였다. 향고래의 대부분은 14 m ~ 16 m의 전장으로 추정되었으며, 9 m ~ 12 m 사이의 개체는 일부지역에서 소수만 존재하였다. 동해에서 발견된 향고래의 전장은 9 m ~ 12 m의 범위에 포함되며, 위도가 높은 뉴질랜드에서 주로 발견되는 성체 수컷 향고래보다는 위도가 비슷한 이오니아해에 서식하는 향고래의 크기와 유사하게 나타나는 것을 확인하였다.
생태학적 연구를 위해 개체의 크기를 측정하고, 그들의 성장을 정량화하는 것은 매우 중요하다. 그러나 고래는 해양이 주요 서식지이고 몸체의 대부분이 수중에 잠겨 있으며, 거대한 몸집으로 인해 측정이 어렵다.[20] 이러한 이유로 과거에는 고래 크기 측정 자료는 대부분 포경으로 죽은 고래에서 획득하였으며, 성장을 정량화하기 위한 연령 및 길이 자료는 포경에 의존하였다.[20,30,31,32] 사진 측정 기법을 이용하여 고래의 길이를 측정하기도 하지만 상당한 시간적 그리고 경제적 투자가 필요하며, 기상상태에 의한 영향을 많이 받는다.[14,33,34,35,36,37] 지속적으로 고래를 따라 이동해야하며 진행방향에서 선박이 나타날 경우 고래가 방향을 바꾸거나 잠수해 버릴 수 있다.[20,38] 그리고 고래의 움직임으로 인해 몸통이 구부러져 있는 경우가 많으며 동일한 지점에서 반복 측정이 어려워 측정 오차가 발생한다.[14,22] 반면에 음향을 이용한 크기 추정은 향고래의 직접적인 조사가 필요하지 않으며, 비교적 간단한 신호처리를 통해 계산할 수 있다. 향고래가 발생시키는 클릭 신호의 IPI와 향고래의 머리 길이, 몸 전체 길이의 관계는 여러 논문들에 의해 측정 및 검증되었다.[9,16,20,39]
국내의 고래 연구는 대부분이 목시 조사에 의해 수행되고 있으며, 아직까지 고래류에 대한 음향학적 연구는 거의 이루어지지 않았다. 최근 들어 일부 고래의 음향학적 특성에 대한 연구가 진행되었지만,[40,41] 개체의 생태학적 특성과 연관시켜 음향 특성을 분석하는 연구는 전무하였다. 본 논문에서 수행한 클릭을 이용한 향고래 전장 추정은 방법론적으로 새로운 것이 아니지만,[20] 음향학적으로 우리나라 근해에 분포하는 향고래의 길이를 추정할 수 있음을 보였고 이는 국내 고래 생태 연구에 있어서 새로운 시도이다. 추후에는 수중청음기 배열을 통해 각 고래의 위치를 분리하고 추적한다면, 음향 신호만으로 향고래의 크기, 분포특성, 이동경로 등을 파악할 수 있으므로 국내 향고래의 생태학적 연구에 유용하게 활용될 수 있을 것이다.
Acknowledgements
본 연구는 국방과학연구소(UD200010DD), 국립수산과학원 수산과학연구사업 고래류 자원 및 생태조사(R2020024)의 지원에 의해 수행되었다.
References
D. W. Rice. "Sperm whale. Physeter macrocephalus Linnaeus, 1758," in
Handbook of Marine Mammals: River Dolphins and the Larger Toothed Whales, edited by S.H. Ridgway and R. J. Harrison (Academic Press, London, 1989).
N. Jaquet, "How spatial and temporal scales influence understanding of sperm whale distribution: A review," Mamm. Rev. 26, 51-65 (1996).
10.1111/j.1365-2907.1996.tb00146.xH. Whitehead, "Sperm whale," in
Encyclopedia of Marine Mammals 3rd ed. edited by B. Würsig, J. G. M. Thewissen, and K. Kovacs (Academic Press, London, 2018).
K.-I. Chang, C.-I. Zhang, C. Park, D.-J. Kang, S.-J. Ju, S.-H. Lee, and M. Wimbush,
Oceanography of the East Sea (Japan Sea) (Springer International Publishing, Switzerland , 2016), pp. 373-387.
10.1007/978-3-319-22720-7F. Caruso, V. Sciacca, G. Bellia, E. De Domenico, G. Larosa, E. Papale, C. Pellegrino, S. Pulvirenti, G. Riccobene, F. Simeone, F. Speziale, S. Viola, and G. Pavan "Size distribution of sperm whales acoustically identified during long term deep-sea monitoring in the ionian sea," PLoS ONE. 10, e0144503 (2015).
10.1371/journal.pone.014450326675588PMC4682957T. W. Cranford, "The sperm whale's nose: Sexual selection on a grand scale?," Mar. Mamm. Sci. 15, 1133-1157 (1999).
10.1111/j.1748-7692.1999.tb00882.xR. Backus and W. Schevill, "Physeter clicks," in
Whales, Dolphins, and Porpoises, edited by K. S. Norris (University of California Press, Berkeley, 1966).
K. S. Norris and G. W. Harvey,
A Theory for the Function of the Spermaceti Organ of the Sperm Whale (Physeter Catodon L) (NASA Special Publication, Washington D.C, 1972), pp. 262-397.
B. Møhl, E. Larsen, and M. Amundin, "Sperm whale size determination: Outlines of an acoustic approach," Mammals in the seas, FAO Series, 3, 327-331 (1981).
B. Møhl, "Sound transmission in the nose of the sperm whale Phy seter catodon. A post mortem study," J. Comp. Physiol[A]. 187, 335-340 (2001).
10.1007/s00359010020511529477B. Møhl, M. Wahlberg, P. T. Madsen, A. Heerfordt, and A. Lund, "The monopulsed nature of sperm whale clicks," J. Acoust. Soc. Am. 114, 1143-1154 (2003).
10.1121/1.158625812942991W. M. X. Zimmer, P. T. Madsen, V. Teloni, M. P. Johnson, and P. L. Tyack, "Off-axis effects on the multipulse structure of sperm whale usual clicks with implications for sound production," J. Acoust. Soc. Am. 118, 3337-3345 (2005).
10.1121/1.208270716334703W. M. X. Zimmer, P. L. Tyack, M. P. Johnson, and P. T. Madsen, "Three-dimensional beam pattern of regular sperm whale clicks confirms bent-horn hypothesis," J. Acoust. Soc. Am. 117, 1473-1485 (2005).
10.1121/1.182850115807035V. Teloni, W. M. X. Zimmer, M. Wahlberg, and P. T. Madsen, "Consistent acoustic size estimation of sperm whales using clicks recorded from unknown aspects," J. Cetacean Res. Manage 9, 127-136 (2007).
H. S. Adler-Fenchel, "Acoustically derived estimate of the size distribution for a sample of sperm whales (Physeter catodon) in the western North Atlantic," Can. J. Fish. Aquat. Sci. 37, 2358-2361 (1980).
10.1139/f80-283J. Gordon, "Evaluation of a method for determining the length of sperm whales (Physeter catodon) from their vocalizations," J. Zool. 224, 301-314 (1991).
10.1111/j.1469-7998.1991.tb04807.xJ. C. Goold, "Signal processing techniques for acoustic measurement of sperm whale body lengths," J. Acoust. Soc. Am. 100, 3431-3441 (1996).
10.1121/1.4169848914321M. Q. Rhinelander and S. M. Dawson, "Measuring sperm whales from their clicks: Stability of interpulse intervals and validation that they indicate whale length," J. Acoust. Soc. Am. 115, 1826-1831 (2004).
10.1121/1.168934615101660L. Rendell and H. Whitehead, "Do sperm whales share coda vocalizations? insights into coda usage from acoustic size measurement," Anim. Behav. 67, 865- 874 (2004).
10.1016/j.anbehav.2003.04.018B. S. Miller, A. Growcott, E. Slooten, and S. M. Dawson, "Acoustically derived growth rates of sperm whales (Physeter macrocephalus) in Kaikoura, New Zealand," J. Acoust. Soc. Am. 134, 2438-2445 (2013).
10.1121/1.481656423968041R. Antunes, L. Rendell, and J. Gordon, "Measuring inter-pulse intervals in sperm whale clicks: Consistency of automatic estimation methods," J. Acoust. Soc. Am. 127, 3239-3247 (2010).
10.1121/1.332750921117772A. Growcott, B, Miller, P. Sirguey, E. Slooten, and S. Dawson, "Measuring body length of male sperm whales from their clicks: the relationship between inter-pulse intervals and photogrammetrically measured lengths," J. Acoust. Soc. Am. 130, 568-573 (2011).
10.1121/1.357845521786922G. Pavan, M. Priano, M. Manghi, and C. Fossati, "Software tools for real-time IPI measurements on sperm whale sounds," Proc. Institute of Acoust. 19, 157-164 (1997).
G. Pavan, C. Fossati, M. Manghi, and M. Priano, "Acoustic measure of body growth in a photo-identified sperm whale," Eur. Res. Cetaceans 12, 254-258 (1999).
G. Pavan, T. J. Hayward, J. F. Borsani, M. Priano, M. Manghi, C. Fossati, and J. Gordon, "Time patterns of sperm whale codas recorded in the Mediterranean Sea 1985-1996," J. Acoust. Soc. Am. 107, 3487-3495 (2000).
10.1121/1.42941910875393V. Drouot, A. Gannier, and J. Goold, "Diving and feeding behaviour of sperm whales (Physeter macrocephalus) in the northwestern Mediterranean Sea," Aquat. Mamm. 30, 419-426 (2004).
10.1578/AM.30.3.2004.419W. M. X. Zimmer,
Passive Acoustic Monitoring of Cetaceans (Cambridge University Press, New York, 2011), 39-85, 119-144.
W. M. X. Zimmer, M. P. Johnson, A. D'Amico, and P. Tyack, "Combining data from a multisensor tag and passive sonar to determine the diving behavior of a sperm whale (Physeter macrocephalus)," IEEE J. Ocean. Engr. 28, 13-28 (2003).
10.1109/JOE.2002.808209G. Giorli and K. T. Goetz, "Acoustically estimated size distribution of sperm whales (Physeter macrocephalus) off the east coast of New Zealand," New Zealand J. Mar. Fresh. Res. 54, 177-188 (2019).
10.1080/00288330.2019.1679843K. Fujino, "On the body proportions of the sperm whales (Physeter catodon)," Whales Res. Inst. Tokyo, Sci. Rep., 1956.
R. Clarke and O. Paliza, "Sperm whales of the southeast Pacific. Part III. Morphometry," Hvalrad Skr, 53, 106 (1972).
T. Kasuya, "Density dependent growth in north pacific sperm whales," Mar. Mamm. Sci. 7, 230-257 (1991).
10.1111/j.1748-7692.1991.tb00100.xS. Cosens and B. Anders, "Size- and age-class segregation of bowhead whales summering in northern Foxe Basin: a photogrammetric analysis," Mar. Mammal Sci. 19, 284-296 (2003).
10.1111/j.1748-7692.2003.tb01109.xS. M. Dawson, C. J. Chessum, P. J. Hunt, and E. Slooten, "An inexpensive, stereophotographic technique to measure sperm whales from small boats," Int. Whal. Commn. Rep., 1995.
J. C. D. Gordon, "A simple photographic technique for measuring the length of whales from boats at sea," Int. Whal. Commn. Rep., 1990.
N. Jaquet, "A simple photogrammetric technique to measure sperm whales at sea," Mar. Mammal Sci. 22, 862-879 (2006).
10.1111/j.1748-7692.2006.00060.xP. J. O. Miller, M. P. Johnson, P. Tyack, and E. A. Terray, "Swimming gaits, passive drag and buoyancy of diving sperm whales Physeter macrocephalus," J. Exp. Biol. 207, 1953-1967 (2004).
10.1242/jeb.0099315107448C. Richter, S. Dawson, and E. Slooten, "Impacts of commercial whale watching on male sperm whales at Kaikoura," New Zealand," Mar. Mamm. Sci. 22, 46- 63 (2006).
10.1111/j.1748-7692.2006.00005.xM. R. Clarke, "Structure and proportions of the spermaceti organ in the sperm whale," J. Mar. Biol. Assoc. 58, 1-17 (1978).
10.1017/S0025315400024371Y. G. Yoon, H. Sohn, K. J. Park, Y.-M. Choi, S. Kim, and J. W. Choi, "Study of Acoustic Characteristics of Common Dolphins Delphinus delphis in the East Sea" (in Korean), Korean J. Fish. Aquat. Sci. 50, 406-412 (2017).
K.-H. Choi, Y. G. Yoon, S. Kim, H. Kim, J. W. Choi, H. S. Bae and K. Park "Analysis of echolocation click signals of Indo-Pacific bottlenose dolphin (Tursiops aduncus) in Jeju Island" (in Korean), J. Acoust. Soc. Kr. 38, 56-65 (2019).


